Medial Olivo-Cochlear (MOC) feedback (mocProc.m)¶
It has now been a well known fact that in the auditory system, an efferent pathway of fibers exists, originating from the auditory neurons in the olivary complex to the outer hair cells [Guinan2006]. This operates as a top-down feedback path, as opposed to the bottom-up peripheral signal transmission towards the brain, affecting the movement of the basilar membrane in response to the input stimulus. The MOC processor mimics this feedback, particularly originating from the medial part of the olivary complex. In Auditory front-end, this feedback is realised by monitoring the output from the ratemap processor which corresponds to the auditory neurons’ firing rate, and by controlling accordingly the nonlinear path gain of the DRNL processor which corresponds to the basilar membrane’s nonlinear operation. This approach is based on the work of [Clark2012], except that the auditory nerve processing model is simplified as the ratemap processor in Auditory front-end.
The input to the MOC processor is the time frame-frequency representation from
the ratemap processor. This is then converted into an attenuation factor per
each frequency channel. The constants for this rate-to-attenuation conversion
are internal parameters of the processor, which can be set in accordance with
various physiological findings such as those of [Liberman1988]. The amplitude
relationship was adopted from the work of [Clark2012]. The time course and
delay of the feedback activity, such as in the work of [Backus2006], can be
approximated by adjusting the leaky integrator time constant rm_decaySec and
the window step size rm_hSizeSec of the ratemap processor.
In addition to this so-called reflexive feedback, realised as a closed-loop
operation, the reflective feedback is realised by means of additional control
parameters that can be modified externally in an open-loop manner. The two
parameters moc_mocIpsi and moc_mocContra are included for this purpose.
Depending on applications, these two can be accessed and adjusted via the
Blackboard system, and applied jointly with the reflexive feedback to the
nonlinear path as the final multiplicative gain factor. Table 30 lists
the parameters for the processor, including the above-mentioned two. The other
two parameters moc_mocThresholdRatedB and moc_mocMaxAttenuationdB are
specified such that the input level- attenuation relationship is fitted best to
the data of [Liberman1988] which is scaled within a range of 0 dB to 40 dB
by [Clark2012].
| Parameter | Default | Description |
|---|---|---|
moc_mocIpsi |
1 |
Ipsilateral MOC feedback factor (0 to 1) |
moc_mocContra |
1 |
Contralateral MOC feedback factor (0 to 1) |
moc_mocThresholdRatedB |
-180 |
Threshold ratemap value for MOC activation in dB |
moc_mocMaxAttenuationdB |
40 |
Maximum possible MOC attenuation in dB |
Fig. 33 shows, firstly on the left panel, the input-output
characteristics of the MOC processor, using on-frequency stimulation from
tones at 520 Hz and 3980 Hz, same as in the work of [Liberman1988]. As mentioned
above, the relationship between the input level and the MOC attenuation
activity through the ratemap representation was derived through curve fitting to
the available data set of [Liberman1988], which is also shown on the plot. An
example of input signal-DRNL output pair at 40 dB input level is shown on the
right panel. The feedback applies an attenuation at the later part of the tone.
These plots can be generated by running the script DEMO_MOC.m.
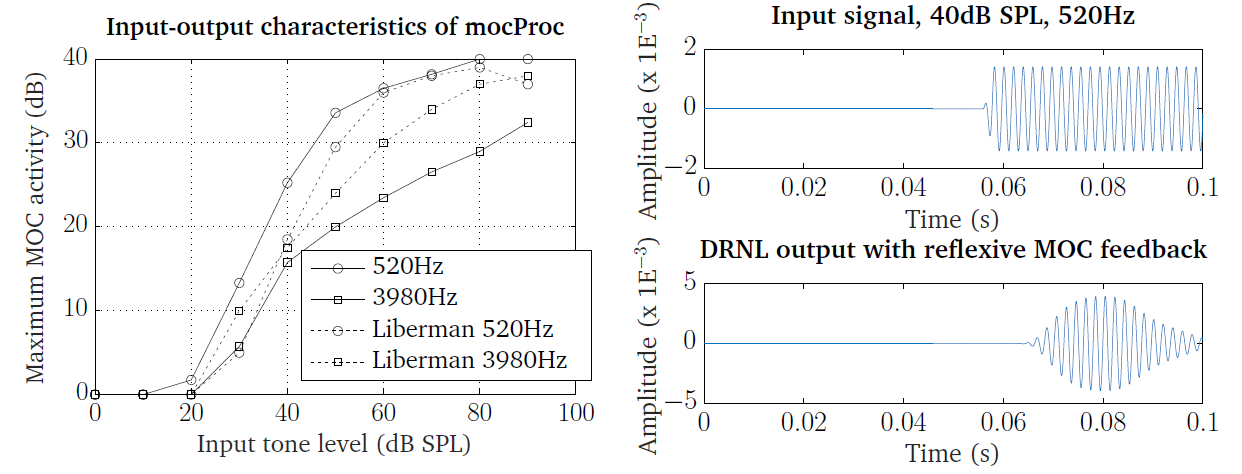
Fig. 33 Left panel: input-output characteristics of the MOC processor for on-frequency tone stimulus at 520 Hz and 3980 Hz. The data set of [Liberman1988], scaled as in the work of [Clark2012], is also shown for comparison. Right panel: DRNL processor output (bottom) from a 50 ms tone at 40 dB SPL and 520 Hz (top), with the reflexive feedback operating.
| [Backus2006] | Backus, B. C. and Guinan, J. J. (2006), “Time-course of the human medial olivocochlear reflex,” The Journal of the Acoustical Society of America 119(5 Pt 1), pp. 2889–2904. |
| [Clark2012] | (1, 2, 3) Clark, N. R., Brown, G. J., Jürgens, T., and Meddis, R. (2012), “A frequency-selective feedback model of auditory efferent suppression and its implications for the recognition of speech in noise.” Journal of the Acoustical Society of America 132(3), pp. 1535–1541. |
| [Guinan2006] | Guinan, J. J. (2006), “Olivocochlear efferents: anatomy, physiology, function, and the measurement of efferent effects in humans.” Ear and hearing 27(6), pp. 589–607, http://www.ncbi.nlm.nih.gov/pubmed/17086072. |
| [Liberman1988] | (1, 2, 3, 4) Liberman, M. C. (1988), “Response properties of cochlear efferent neurons: monaural vs. binaural stimulation and the effects of noise,” Journal of Neurophysiology 60(5), pp. 1779–1798, http://jn.physiology.org/content/60/5/1779. |